I found that TGF Alpha is highly correlated with the so-called Thymidine described as a synchronization element during genomic processes. I see synchronization as a possibility of migration of the correct behaviour, i.e., hair growth and phases cycling, anagaen–>catagen–>telogen–>exogen–>early Anagen, practised by the healthy tissue towards the closest bald spots. In this model the researcher could gain-cumulate extra small squares of healed tissue in.
The curcumin although has proved itself in terms of carcinogenic growth inhibition and anti-inflammation as well, which is the main fundamental approach to face PB, should some how slow/hinder at some point the migration of non-pathogenic behaviour, hair, DPs, etc…, phases transition, etc.
I am missing some sources, google results seem unstable I think but,
I found that a main pathway for TFG Alpha alpha growth factor is through inhibition of Tyorsine Kinase
AND
That curcumin inhibits tyrosine kinase
Thus curcumin should stop/hinder genomic synchronization, consequently, in a post terminal phase of PB, AGA, etc will not help a lot with hair, hair-cycling moving back to bald spots.
If you consider that Geftinib is a Tyrosine Kinase inhibitor and that Curcumin is taken instead of the industrialized drug in case of Gefitinib-resistant NSCLC
If you consider that
“Current studies describe that curcumin can inhibit initiation, course and metastasis of various tumor lines. These effects are related to the negative regulation of various transcription factors (NFκβ, AP-1, Egr-1, STATs, PPAR-gamma, nrf2, EpRE, p53, CBP, CREB-BP, β-catenin, HIF-1, Nrf −2, PPAR-γ, ERE) [[51], [52], [53]], growth factors (FGF, HGF, EGF, PDGF, TGF-β1, VEGF) [51], cell proliferation factors (cyclin D1, c-myc), cytokines, protein kinases (JNK, Akt, AMPK) [54] and many other cell signaling proteins [51] including COX2, 5-LOX and tubulin. Due to the fact that the curcumin molecule exerts an effect on many different signaling pathways, the probability of resistance to this compound is lower [54], on the other hand, there are great difficulties in controlling and directing the action of natural pleiotropic particles.”
You will get to accept that curcumin wont be the medicine for a full head of hair therapy.
But stills one of the best in terms of depth-medicine
Among all spices, turmeric (Curcuma longa) has been proven for its better anticancer potential. In this review different molecular mechanisms including cell cycle arrest; G0/G1 and/or G2/M phase cell cycle arrest by up-regulating Cdk inhibitor, p21/WAF/CIPI and p53, inhibition of transcriptional factors; NFκB, AP-1, TNFα, IL, STAT-3, and PPAR-γ, downstream gene regulation; c-myc, Bcl-2, COX-2, NOS, Cyclin D1, TNFα, interleukins and MMP-9, growth factors; bFGF, EGF, GCSF, IL-8, PDGF, TGFα, TNF, VEGF and cell adhesion molecules; fibronectin, vitronectin, and collagen which are involved in angiogenesis and metastasis
I am searching the TGF alpha for Tea and the transcription factors. Please assist.
In this study, [6]-gingerol was demonstrated to inhibit both the proliferation and invasion of AH109A cells in vitro (Fig. 1) . [6]-Gingerol concentrations that could significantly suppress the proliferation of AH109A cells (Fig. 1a, 12.50–200 μM) were lower than those which could suppress the invasion of AH109A cells (Fig. 1b, 50–200 μM). [6]-Gingerol commenced to significantly inhibit the hepatoma proliferation at a concentration of 12.5 μM and almost completely inhibited (6% of control) at 200 μM (Fig. 1a), whereas it significantly inhibited the invasion at 50 μM, which was four times as high as 12.5 μM, and the highest inhibition observed at 200 μM was 34% of control (Fig. 1b). These results suggest that the inhibition of proliferation by [6]-gingerol might not be a main cause of its inhibitory action against the invasion.
To elucidate the mechanisms of inhibition of hepatoma cell proliferation by [6]-gingerol, the effect of [6]-gingerol treatment on cell cycle and on the rate of apoptosis was examined by using flow cytometry. [6]-Gingerol was found to suppress proliferation from lower concentrations (25 μM∼) by increasing doubling time of AH109A cells through accumulation in the S phase (Fig. 2). Further, [6]-gingerol increased the rate of apoptosis at higher concentrations (100–200 μM) (Fig. 3). [6]-Gingerol has been reported to induce apoptosis in HL60 cells (Lee and Surh 1998). It is apparent that different cell lines have different sensitivities to [6]-gingerol but the primary target in cell cycle and apoptosis may be the same in these cell lines. Although the doses of [6]-gingerol which induced apoptosis were higher than those which induced cell cycle arrest in AH109A hepatoma cells, these results suggest that [6]-gingerol affects both cell proliferation and cell death which at least partly account for the inhibitory effect of [6]-gingerol on the proliferation of AH109A cells.
Our previous works have demonstrated that the invasion of AH109A cells is accelerated by ROS (Kozuki et al. 2000). In the present study, we therefore examined the effect of [6]-gingerol on the ROS-potentiated invasive activity using both HX–XO system and H2O2. [6]-Gingerol was found to inhibit the ROS-induced elevation of the AH109A invasion (Fig. 4). [6]-Gingerol was also found to scavenge intracellular peroxides (Fig. 5). We have found that ROS can induce gene expression of hepatocyte growth factor (HGF), which is known as a cell motility factor (Parr and Jiang 2001), in M-cells as well as AH109A cells (Miura Y et al. 2003b). Thus, HGF produced by both AH109A and M-cells may potentiate the motility of AH109A cells and also may induce the retraction of M-cells, leading to acceleration of the AH109A invasion. Provided that [6]-gingerol, like a polyphenol resveratrol (Miura D et al. 2004), suppresses the production of HGF through its anti-oxidative activity, [6]-gingerol may diminish the induction of the retraction of M-cells as well as the motility of AH109A cells, this leading to the effective reduction of the AH109A invasion by reducing the functions of both cells at the same time. Since prostaglandins are shown to enhance the invasion of hepatoma cells (Miura D et al. 2003), a possibility that [6]-gingerol, a cyclooxygenase inhibitor (Kim et al. 2004), interrupts prostaglandin synthesis and hence inhibits the invasion cannot be ruled out. Lee et al. have recently reported that [6]-gingerol inhibits cell adhesion, invasion, motility and activities of matrix metalloproteinase (MMP)-2 and 9 in MDA-MB-231 human breast cancer cells (Lee et al. 2007). AH109A cells are different from this breast cancer cells, for instance, in the productivity of MMPs; AH109A cells do not produce MMPs. Thus, different action sites as well as common action sites of [6]-gingerol may be existent between these two cancer cells. Further studies are required to clarify these aspects.
In summary, we clearly demonstrated that [6]-gingerol inhibited the proliferation and invasion of AH109A hepatoma cells in culture, the proliferation being more strongly suppressed than was the invasion. [6]-Gingerol induced cell cycle arrest at lower concentrations and apoptosis at higher concentrations in the hepatoma cells. It also suppressed the ROS-induced increases in invasive capacity and intracellular peroxide levels. These results suggest that [6]-gingerol affects both cell proliferation and cell death that account for, at least partly, the inhibitory effect of [6]-gingerol on the proliferation of AH109A cells. They also suggest that the anti-oxidative property of [6]-gingerol may be involved in its anti-invasive action. [6]-Gingerol may have promising beneficial effects in preventing tumor growth and metastasis and may be of significance from the aspect of nutritional control of cancers.
At least, I could consider ginger for the amelioration of the signaletic environment, that was the reason I developed this Behaviour migration theory in the first place.
Reservatrol extracts such as Grape Vinaigre or Wine may be a medicine targeting Dermall papilla mobility or behaviour migration, differentiation propagation, trigger-induction behaviour behaviour mobility and transcription in target close tissue etc.
Black chocolate extract as well,
Sources
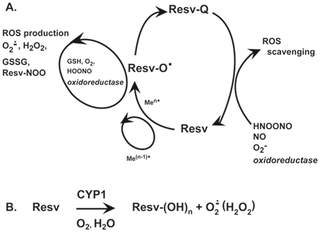
Fig 3.
Resveratrol increases FOXO3 transcriptional activity and expression of FOXO3 target genes.
The size of a hair follicle is thought to be determined by the volume of its dermal papilla. The volume of the dermal papilla depends on the number of cells it contains and on the volume of the extracellular matrix. To establish which of these two variables is related to differences in hair follicle size we performed a stereologic study on 235 hair follicles from different sites, including male facial skin (beard), female facial skin, and scalp. In facial follicles there was a strong correlation between the area of the hair cortex and the volume of the dermal papilla. The area of the hair cortex also correlated with the number of cells in the dermal papilla and with the volume of dermal papilla per cell. In scalp hair follicles, where there was a smaller range of sizes, the correlations between these variables were weaker. In large male facial follicles the mean total dermal papilla volume was almost 40-fold higher than in vellus follicles from female facial skin. This difference was associated with a mean 17-fold greater number of cells in the dermal papilla and a 2.4-fold greater volume associated with each cell. Intermediate results were obtained in scalp follicles. In many regions of the skin hair follicles enlarge in response to androgens during adult life hair. Our results imply that the increase in the volume of the dermal papilla in these follicles is due to an increase in the number of cells, either through proliferation or through the migration of cells from the follicular dermal sheath, and to an increase in the amount of extracellular matrix per cell. As androgens are thought to act primarily on the dermal papilla, these changes may have a direct bearing on the mechanism of androgen-mediated alterations in hair follicle size.
This is an advertising site for paid
advertisers to showcase successful hair restoration results only. It is not the
mandate of this site to engage in the discussion of failed, unsuccessful
procedures, lawsuits, litigations, refunds or complaint cases. Surgical hair
restoration procedures carry risks. Please do thorough research, consult your
own physician and investigate a doctor's background carefully before making a
decision. By proceeding to use our site, you agree to abide by our Terms of Use & Privacy Policy at http://hairsite.com/terms-of-use/ where you can also find a list of HairSite's sponsoring physicians.